

I recently stumbled upon a copy of The Dinosaur Heresies, by Robert Bakker, published in 1986, for $6 in as-is condition at the local used book store. I’d heard of Bakker and his book before–he’s a preeminent paleontologist and one of the instigators of the Dinosaur Renaissance of the ’80s and ’90s–but I hadn’t thought it would be worth going out of my way to obtain because it’s so old, and more modern dinosaur books like Dinosaurs Rediscovered (Benton) and The Rise and Fall of the Dinosaurs (Brusatte) would do the same job but better. Boy was I wrong–The Dinosaur Heresies is an amazing book. It’s full of well-thought-out, evidence-backed hypotheses about how dinosaurs would have lived, moved, and behaved, and countless beautiful (and sometimes satirical) illustrations by the author that still hold up to modern scrutiny. In this post, I’ll go over some of the more interesting things I got out of reading this book, and discuss what else is known about them now.
Numerical-Ecological Clues to Dinosaur Habits
A major point of this book was presenting the overwhelming evidence that dinosaurs were in fact warm-blooded, since at the time of writing this wasn’t widely accepted. As a modern reader though, the thing that struck me is just how much you can infer just from fossil occurrence rate. Bakker used quite a few numerical-ecological indicators to show that dinosaurs were probably warm-blooded and active, including modern-day niche divisions between warm- and cold-blooded creatures, predator-prey ratios, and genus lifetime.
In the modern world, cold-blooded tetrapod (non-fish) species vastly outnumber warm-blooded ones in small, land-living roles (6000 reptile and amphibian species to 2400 mammal species), small freshwater roles (1290 reptiles and amphibians to 50 mammals), and large freshwater roles (15 reptiles to 1 mammal, the hippo). But warm-blooded species vastly outnumber cold-blooded ones as large land animals (100 mammals to 5 reptiles), which is what dinosaurs were–large land animals. Thus, this implies that dinosaurs were more likely analogous in function to modern large mammals than to oversized lizards.
Warm-blooded animals are much less energy-efficient than cold-blooded ones, since we’re always wasting heat to the environment and spending energy moving about frenetically instead of basking. So, as I’ve written about in a previous post, warm-blooded animals require around ten times as much food as cold-blooded animals of the same weight. This means that for carnivorous species, you’d expect prey to outnumber predators by a lot more if the predators are warm-blooded than if they’re cold-blooded, and indeed this is borne out by modern examples. Lions, hyenas, and cheetahs are outnumbered 99 to one by their prey on the African savanna, while the wolf spider, a cold-blooded predator, is only outnumbered three to one. We see this pattern in the fossil record as well: Dimetrodon, the famous Early Permian finbacked apex predator, which was almost certainly cold-blooded, is quite abundant in its habitat, with a similar ratio to its prey as the wolf spider.

Having established this pattern, Bakker then looks at environments in which we don’t know whether the predators were warm- or cold-blooded, and what he finds makes a lot of sense. The Late Permian proto-mammals and Early Triassic erythrosuchids (terrestrial croc relatives) have predator-prey ratios intermediate between that of the finbacks and spiders and that of modern mammals, being outnumbered by prey about nine to one, indicating that they may have been mesotherms, with metabolisms intermediate between fully warm- and fully cold-blooded predators. And predatory dinosaurs were outnumbered about thirty to one on average through the Jurassic and Cretaceous, indicating they had warmer blood than those that came before.
But why did predatory dinosaurs enjoy a higher ratio than modern predators? Were they colder-blooded than lions and hyenas? No–mammalian predators in the Eocene Epoch, 56 to 34 million years ago, had a similar predator-prey ratio as the dinosaurs, and these mammals were as warm-blooded as modern mammals. It’s just that in the modern world, predators are persecuted more than prey, and don’t fill out the environment as much as they could in the absence of humans.
One more unexpected place to find evidence of warm-bloodedness in the fossil record is in the lifetime of genera. In warm-blooded animals, the turnover rate, the time between the appearance of a genus and its extinction, is much quicker than in cold-blooded animals. Across the Cenozoic Era, mammal genera only last 3 to 5 million years on average before they’re replaced by a new one, while croc and turtle genera last 30 to 40 million years. And in places where millions of years of dinosaur history is recorded, we see a similar rate of turnover as we do in mammals, with Chasmosaurus being replaced by Anchiceratops and then Triceratops after a reign of less than 5 million years each, for example.
Biomechanics: Stegosaurus
Another of the cool things in this book are Bakker’s detailed derivations from first principles of dinosaur behaviors, postures, and abilities. He looks at all aspects of the skeleton–joint orientation and construction, bone texture, muscle attachment point and lever length, modern analogues, etc–and comes up with a convincing picture of what a dinosaur would’ve looked and acted like. The one he spends the most time on across multiple chapters of the book is Stegosaurus, the plate-backed, thagomizer-tailed, tiny-headed plant eater from the Late Jurassic United States.
To start with, Stegosaurus’s hind legs are much longer than its front legs. Old-school dinosaur orthodox adherents apparently said that “it was just a clumsy dinosaur” and “it couldn’t walk too fast or else the back legs would overtake the front”. But obviously this arrangement must have been adaptive, or else it wouldn’t’ve evolved. What could it be for?
Bakker proposes that the large rear end and heavy tail would’ve put Stegosaurus’s center of gravity just forward of its hips, allowing it to easily pivot its body about the hind legs. The ability to pivot agilely would be invaluable if your main deadly weapon is your tail, which you would want to keep facing your enemy at all times. Stegosaurus also had massive attachment points for its shoulder muscles, which it would’ve used to push its upper body sideways to initiate those pivots.
Another feature of the aftward center of gravity is the ability to tip up into a tripod stance, for browsing on high-up foliage. While more basal, bipedal dinosaurs had stiffened tails to help with balance while running, Stegosaurus ditched the stiffness in favor of a very flexible tail, which would’ve been able to bend where it contacted the ground (and allow more precise and whippy strikes with the thagomizer). Stegosaurus also had tall vertebral spines over the hips, which provided leverage for tendons that acted like the bracing cable on a crane, allowing the animal to hold up its chest and head at an angle.
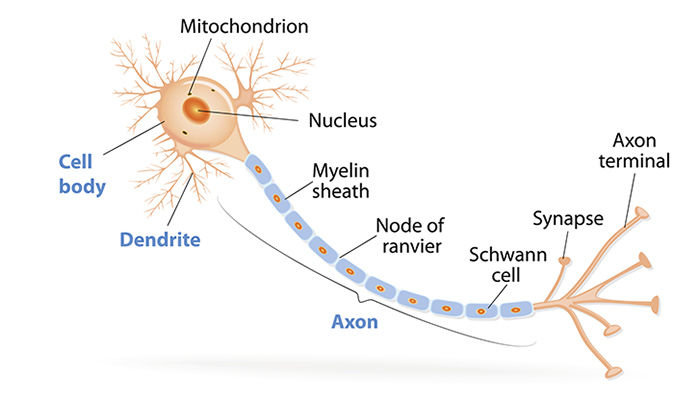
One thing that suprised me was that the second brain hypothesis actually has some basis in reality! It’s often brought up, when the topic turns to wacky old theories about dinosaurs, that nineteenth-century paleontologists suggested that since Stegosaurus’s head was so small and its body was so large, it needed a second brain, which was housed in its hips, to control the hindlimbs and tail. Obviously, no known tetrapod has a second brain, but this is probably the most ungenerous statement of the hypothesis possible. The reason they thought there might be a brain in the hips is because there is indeed a very large cavity, thirty times larger than the braincase, inside the vertebrae above the hips, indicating that the spinal cord swelled there to a much larger diameter than elsewhere. And this lumbosacral expansion, as it’s called, is actually present in all tetrapods, including you! Animals’ spinal cords are not uniform in diameter–they’re larger in the upper back, where lots of nerves exit to control the arms, and in the lower back, where lots of nerves exit to control the legs. In terms of pure cell volume, this makes sense. I’m sure you’ve seen a picture of a neuron before, but in case you haven’t, here’s one:
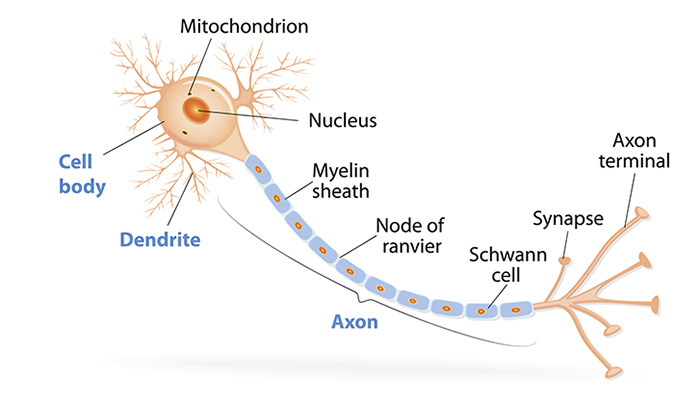
The ball-shaped orange part on the left is the cell body, and literally all of these live inside your brain and spinal cord. The skinny part on the right, the axon, extends out from your spinal cord, all the way to your furthest extremity. Yes, a single cell (well, many of these uber-long cells in parallel) runs the entire length from your tailbone to your toes. And if a lot of these are needed in one area, the cell bodies will be a bit more crowded, making the spinal cord larger in cross-section in that area. In animals that are very one-limbset-dominated, such as an ostrich, which has enormous legs and puny arms, the lumbosacral expansion is quite impressive, with double the cross-sectional area of the rest of the spinal cord. Since Stegosaurus had such large hind legs and such an important tail, it’s no wonder that its lumbosacral expansion was record-setting.
(Note: birds’ lumbosacral expansions are accentuated further by the presence of an organ known as a glycogen body, which resides in the center of the spinal cord over the hips. It’s unknown what the glycogen body does, other than hold lots of glycogen, but it’s probable that Stegosaurus had one as well, making its lumbosacral expansion look even more extreme.)
I’ve saved the most controversial thing about Stegosaurus for last: the function of the plates. It’s still not fully agreed upon what the plates were for; the three hypotheses are (a) sexual display and intimidation, (b) thermoregulation, like an elephant’s or rabbit’s ears, and (c) defense. Of course, these functions are not mutually exclusive, and in all likelihood they served all three purposes to at least some extent. But Bakker argues that their main purpose was active defense, and that Stegosaurus would have been able to flap the plates toward the threat to protect its flanks. He cites the plates’ light construction as helping the animal to move them easily, and their rugose (roughened) texture on the bottom third of the plate as muscle attachment points, and proposes that the top part of the plate would’ve been covered in a keratin (horn) sheath that could’ve given them a wickedly sharp edge. However, these arguments could easily be drawn the other way: the light construction hinders their usefulness as armor, since they would be fragile, and the bottom third was probably anchored deeply in the skin, preventing them from achieving a hinging movement. Furthermore, a hollow pipe structure is present in the bottom of the plate, which probably housed a large artery. This branches into smaller pipelets that would have supplied blood to the rest of the plate, supporting the idea that the plates were important for thermoregulation and display (flushing them with blood would’ve given them an attractive red or pink color).
A paper studying the histology of the plates, published in 1986, the same year as The Dinosaur Heresies, proposes some falsifiable predictions we should expect if the plates were carried vertically, horizontally, or if they were flappable. If they were vertical, we should expect mirror-image attachment marks on both sides of the plate; if they were horizontal, we should see directional attachment marks; and if they were flappable we should see much deeper attachment marks on the medial side where strong muscles would have pulled them upright, and shallower ones on the distal side, which would have been assisted by gravity.
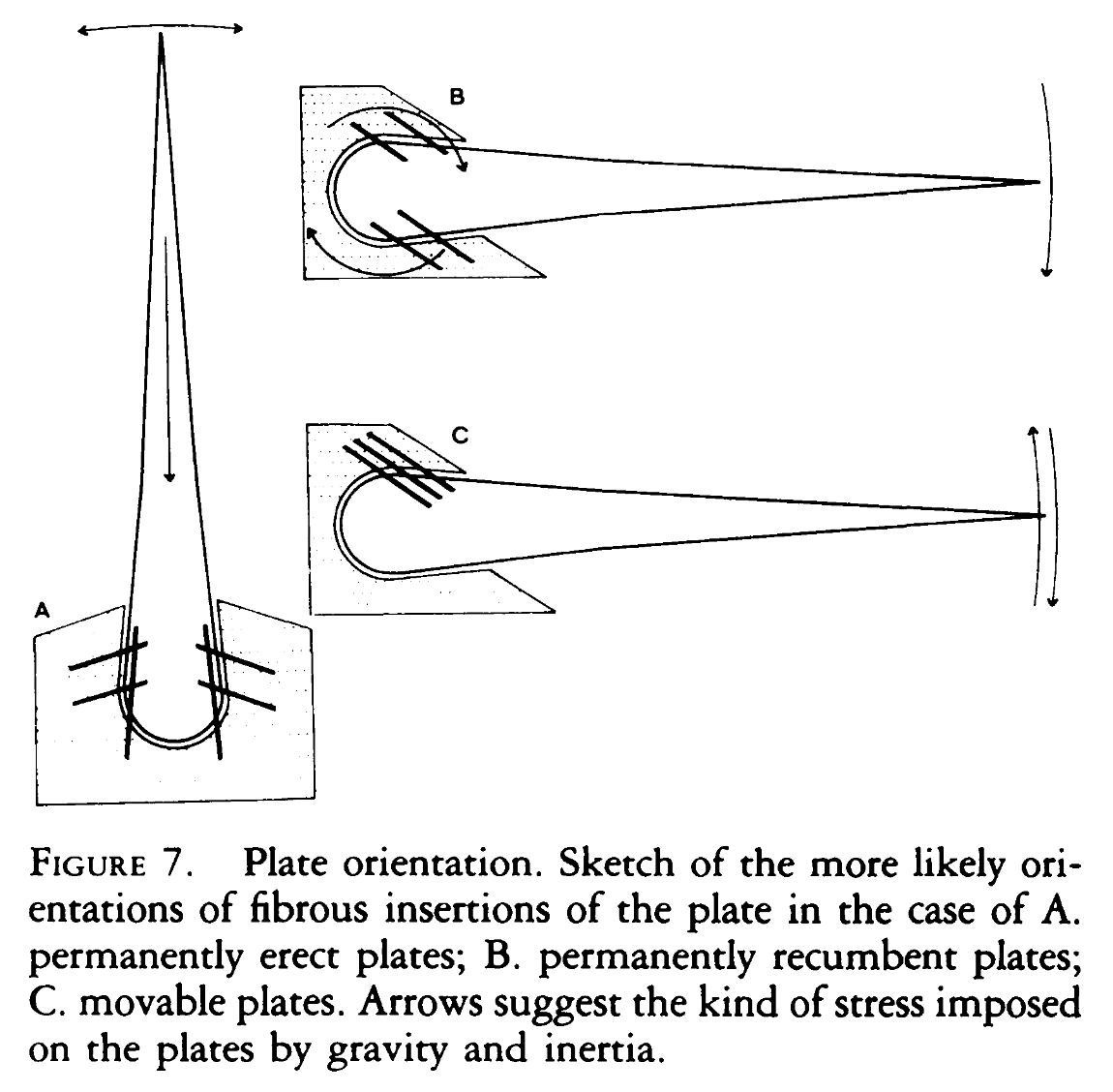
In reality, we see something most similar to A, indicating that the plates were probably buried vertically and mostly immovably in the skin. An additional prediction we would expect to find if the plates were used as defense is the presence of injuries–plates that were wounded and then healed. While we’ve found quite a few Stegosaurus plates, many of which are damaged or fragmentary, none have shown signs of healing, meaning they were probably damaged after death. We have found two injured thagomizer spikes though, indicating that these were indeed used as weapons.

Biomechanics: Triceratops
Another dinosaur Bakker spends a lot of time discussing is the famous horned behemoth from Latest Cretaceous North America, Triceratops. It’s long been debated what these dinosaurs’ neck frills were for–display, defense, “species recognition”, or something else?–and, like Stegosaurus’s spikes, it’s likely that they fulfilled all these functions to some extent. But Bakker proposes a couple of additional uses for the frill that I hadn’t heard of before, and which make a lot of sense. First, ceratopsians’ jaw muscles actually passed through holes in the back of their heads and anchored to the front of the frill, giving them the strongest bite of any herbivore. However, these muscles didn’t extend very far up the frill, so the rest of it must’ve served some other purpose.
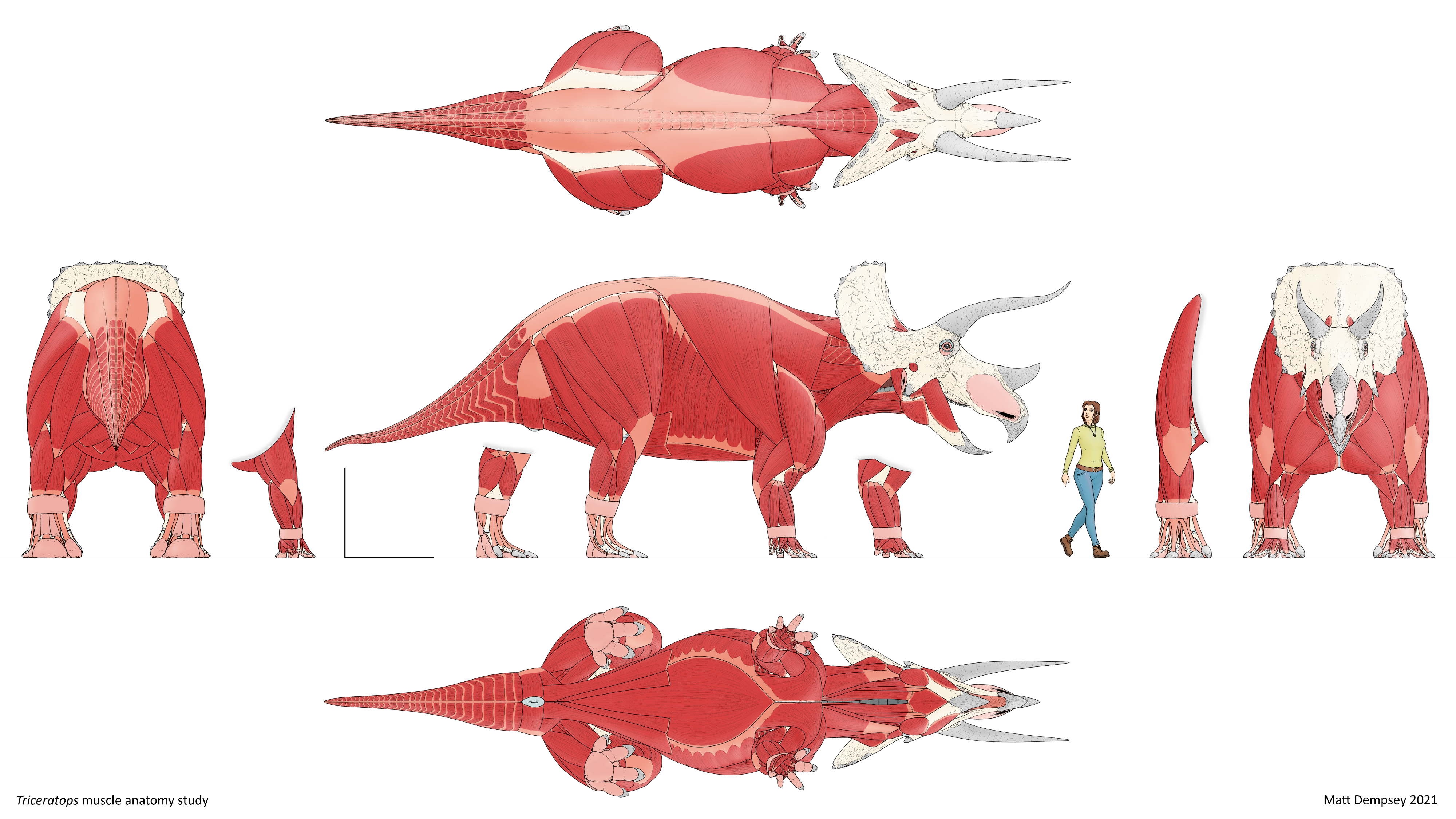
Another biomechanical function Bakker proposes for the frill is to counterbalance the long, heavy snout and horns, allowing Triceratops to toss and swivel its head quickly and easily. In fact, the place where the skull joins the neck is nearly directly under the center of mass of the whole skull. So, in addition to the obvious display and defense functions, the frill would have served a couple of very important biomechanical purposes in the animal’s day to day life.

Bakker also draws comparisons between Triceratops and modern large animals such as rhinos and elephants, to make guesses about how the dinosaur would have moved. Rhinos are good gallopers, able to run 56 kilometers per hour (35 miles per hour), while elephants aren’t able to completely leave the ground and are only capable of a 35 kph (22 mph) speedwalk. You can predict the difference by looking at their skeletons: rhinos have short, sturdy bones with joints designed to be held at an athletic low angle, while elephants have long, thin bones that are held nearly straight like a column. Rhinos also have impressively long shoulder blades, which anchor the muscles used to power their front legs. Triceratops was more similar to a rhino, with thick, angled leg bones, and super-elongated shoulder blades, and thus can be expected to have been capable of high speeds. It also concentrated more of its mass in its legs, making it probably faster than a rhino of the same overall weight. Sauropod (long-necked) dinosaurs like Camarasaurus had longer, thinner, more columnar bones like an elephant, and probably could not jump or gallop, but they also had long shoulder blades, giving their front legs a powerful driving stroke.
Things the Scientific Establishment Could’ve Accepted Earlier
Something else that impressed me a lot about this book is that Bakker got a lot of things right that would not become common knowledge until much, much later–like, thirty years later. Here’s an abbreviated list:
Tyrannosaur lips. While Jurassic Park was inspired by a lot of the work coming out of the Dinosaur Renaissance, including this book, it depicted its Tyrannosaurus as lipless, alligator-style, with teeth that show even when the mouth is closed. But osteological correlates (bone texture) on dinosaur skulls indicates that they had lizard-like lips, a fact that Bakker highlights and also adheres to in his illustrations.
Lagosuchus as dinosaur-pterosaur ancestor. The family tree of dinosaurs, pterosaurs (flying reptiles), and pseudosuchians (croc relatives) has gotten shaken up many times before, during, and after the Dinosaur Renaissance. Bakker doesn’t have it exactly correct in this book (more on that in the next section), but he does hit one thing right on the head: dinosaurs and pterosaurs are closely related, and they descended from a common ancestor that looked something like Lagosuchus, a small, agile, warm-blooded Triassic archosaur.
Dinosaurs invented flowers. This one is more of a pet theory of Bakker’s, but I’m a fan. Flowering plants, called angiosperms, which make up a vast majority of plant species today, first appeared in the Early Cretaceous as unassuming water-plants, but didn’t take over the world until the Late Cretaceous. People often point to coevolution with pollinators as the factor that took some time to really bear fruit (pun intended), but Bakker argues that dinosaurs had an equally important role in the rise of angiosperms. Herbivorous dinosaurs in the Late Jurassic were nearly all high browsers, with tons of long-necked sauropods and tripod-able stegosaurs coexisting in the same environment. I’ve discussed how the niches were partitioned in a previous post. These herbivores would’ve mostly ignored young sprouts and focused on mature vegetation, selecting for trees that didn’t have to grow quickly, but had to get tall: conifers are the masters of that evolutionary strategy. But the Jurassic ended with a minor extinction event that killed off the stegosaurs and decimated the diversity of the sauropods, and allowed new types of herbivores to evolve: hadrosaurs (duck-billed dinosaurs), ceratopsians (horned dinosaurs), and ankylosaurs (tail club dinosaurs), which all held their heads close to the ground. Now, the selection pressure on plants was to grow fast and reproduce early, or else you’d be literally nipped in the bud by a big chomping beak. This may have given angiosperms the foothold they needed to spread and diversify.
But why did all the high browsers exist at the same time, and then all the low browsers? Doesn’t that leave open niches for the opposite style of herbivore, which we should expect to be full because nature abhors a vacuum? Well, it’s not fully understood. But in actuality, markets are never perfectly efficient, and unlike day traders, animals can’t evolve with a goal in mind–evolution will never select for something detrimental now in preparation for a payout later. So if all the successful herbivores happen to come from one or two groups, and those groups are both good at the same type of thing, competition will drive them to become better at that thing, rather than completely changing gears to do a different thing, since they’d have to pass through a maladapted intermediate stage first. Furthermore, there were actually some Jurassic low-browsers, like the little bipedal ornithischian Nanosaurus (also known as Drinker, Othnielia, and Othnielosaurus), and there were some Cretaceous high-browsers, like the giant sauropods Argentinosaurus and Alamosaurus. And, there were additional factors influencing the success of different dinosaur groups, like climate and geography (sauropods kept away from swamps, mountain raising created deserts or prairies where there used to be tall forests, land bridges allowed dinosaurs to disperse or sea formation kept populations apart, etc.).
Things Bakker Was Wrong About
Of course, no one is perfect, and you can expect lots of what we now know to be errors in a science book from 1986, as science has marched onward since then. Here’s an impressively brief list of things Bakker got wrong.
Ceratosaurus cranial kinesis. Ceratosaurus was a primitive large predatory dinosaur that had a very lightly built skull and jaw. Bakker proposes that it may have been able to widen its skull, sort of like a snake, to swallow larger things than it would otherwise have been able. While it’s true that modern lizards and birds exhibit a lot of cranial kinesis (you can see a parrot’s skull in motion in this video), non-avian dinosaurs lacked the linkages and musculature required to voluntarily move their skulls. The fact that some of them had what appear to be joints in their skulls is probably just a plesiomorphy, or characteristic inherited from their ancestors, that isn’t used anymore.

Anti asteroid. While the Alvarez hypothesis, which proposes that a large extraterrestrial object impacted the earth at the Cretaceous-Paleogene boundary and that this caused the mass extinction that killed the non-avian dinosaurs, was widely known in 1986, it wasn’t until 1990 that scientists uncovered the smoking gun–the enormous Chicxulub crater that matched up with the layer of extraterrestrial iridium in K-Pg rocks. Bakker agrees that an asteroid probably did hit the earth at the K-Pg boundary, and that it may have finished off the dinosaurs, but he contends that environments were already struggling at that point due to climatic factors: global cooling, and the faunal interchanges enabled by receding seas. However, in addition to the discovery of the crater, more recent statistical studies of fossil occurence (and more fossil discoveries that have taken place since 1986) show that dinosaur diversity had not actually been lower until the extinction event.
Phytodinosauria. While Bakker’s dinosaur family tree proposed in the book isn’t quite right, it’s actually an admirable effort, given the undetailed “hub and spoke” model that prevailed at the time. When completely rewriting a phylogeny, one can expect some errors on the first go. One of those is Bakker’s grouping of the sauropods and ornithischians together in a group called Phytodinosauria (“plant dinosaurs”). It’s now thought that sauropods either belong alongside the theropods (which includes most of the meat-eating, bipedal dinosaurs, as well as birds), or in their own group with the herrerasaurs (a group of primitive meat-eating dinosaurs).
Pterosaurs are dinosaurs. This is more of a case of changing definitions rather than incorrectness. We now define the group Dinosauria as the last common ancestor of the house sparrow, Triceratops, and Diplodocus, and all its descendants, and on Bakker’s tree this group is still a proper clade; it’s just not given a name. He employs a more inclusive definition of Dinosauria, as the last common ancestor of the house sparrow and Quetzalcoatlus, and all its descendants, which is the group we now define as Ornithodira. So, pterosaurs are not dinosaurs, but only by fiat.
Sauropod live birth. This isn’t discussed in much detail, but is mentioned as a side note when discussing dinosaur growth rates and the immense size difference between hatchlings and adults. He believed sauropods may have been viviparous because (a) no sauropod specimens smaller than 230 kilograms had been found, and that’s way too large to have hatched from an egg, (b) the sauropod pelvic canal is quite wide, and (c) if they did hatch from eggs, the babies would have been so tiny compared to the adults that they would have been crushed underfoot, or at least unable to keep up with the herd. However, since the writing of the book, we’ve discovered many unequivocal specimens of sauropod eggs and nests, some of which have embryos still inside (which we know via CT scan). So, (a) is now irrelevant. (b) is probably the case just because sauropods themselves were very wide, and there was no reason for the pelvic canal to be unnecessarily restricted. And as for (c), we now know based on fossil trackways (footprints) that sauropods self-segregated by age: babies would have formed all-baby herds, or “creches”, and lived independently, and quite differently in terms of ecology, than the adults.
Sauropod nostril location. Sauropod nares, the nose openings in their skulls, are positioned high up on the forehead, so older art, Bakker included, depicts the external nostrils as being there as well. However, we now believe that long, soft-tissue nose channels would have served a cooling function for these enormous, hot-blooded beasts, and the nostrils would’ve been on the end of the snout.
Eight-spiked Stegosaurus ungulatus. A particular subspecies of Stegosaurus was initially published as having had eight thagomizer spikes, but four of those were later realized to have belonged to a different individual.
Conclusion
Overall, I would highly recommend this book. The illustrations alone are really inspiring and useful–I’m definitely going to be using some of his skeletal top views as reference in the future–and the writing style is dynamic, entertaining, and accessible. It’s probably more inspiring even than the aforementioned Dinosaurs Rediscovered, which is more dry and technical, and The Rise and Fall of the Dinosaurs, which is a higher-level overview and doesn’t bring the animals to life as well, both of which are also very much worth the read. And no, I didn’t just summarize the entire book. There’s a lot more good stuff to be learned I didn’t touch on at all, like the debunking of mass homeothermy, the biomechanics of hadrosaurs, what headbutting structures say about the rest of an animal’s lifestyle, and the noisemaking structures dinosaurs possessed.
The Dinosaur Heresies is NOT TO BE CONFUSED WITH The Pterosaur Heresies, a crackpot blog run by Dave Peters. He also runs Reptile Evolution [dot] com. Both of these are full of scientific-looking diagrams and crazy theories with no grounding in reality.
Image Credits
Dimetrodon Neuron Stegosaurus Triceratops muscle study Lagosuchus Ceratosaurus
{kind=link}